2. Energiatermelés az emberi szervezetben
2.1. Honnan származik az energia?
Életünkben minden tevékenység alapja az annak végzéséhez szükséges megfelelő mennyiségű energia előállítása. Mivel a sportolás egy felfokozott mozgással járó tevékenység, az energiatermelés folyamatának megértése és befolyásoló tényezőinek ismerete központi szerepet kap a sportélettanban. Az energia előállítása során lényegében nem teszünk mást, mint hogy a tápláléklánc elején helyet foglaló autotrof élőlények (zömmel növények) által különböző molekulák kémiai kötéseibe bezárt napenergiát elérhetővé tesszük saját magunk számára.
A zöld növények egyszerű, szervetlen anyagokból (CO2, H2O) és fényenergiából állítják elő az élet alapját jelentő szerves nagymolekulákat, miközben oxigént adnak le a környezetükbe melléktermékként. A szerves anyagok felépítése első lépésben szénhidrátokat (glükóz; C6H12O6) jelent, majd ezeknek különféle biokémiai módosításával állítják elő a lipideket (zsírok, olajok), a fehérjéket és a nukleotidokat. Biológiai tanulmányainkból emlékezhetünk, hogy mindenféle szervesanyag („biomolekula”) gerincét az összekapcsolódó szénatomokból álló szénváz alkotja, és ezen helyezkednek el az anyag típusától függően a hidrogén, az oxigén, nitrogén, a foszfor és a kénatomok (valamint egyéb atomok és ionok az előző hathoz képest sokkal kisebb arányban).
A fotoszintézis folyamata tehát egyszerűen (és molnyi mennyiségekre nem rendezetten) így néz ki:

A növények (és egyéb autotrof élőlények) tehát előállítják az élő szervezetek számára szükséges nagyobb molekulákat. Ezeket mi a táplálékkal felvesszük, és az emésztés folyamatában kisebb egységeikre bontjuk. Ezeket a kisebb egységeket, úgy mint
- a szénhidrátbontásból származó glükózt,
- a lipidek alkotóit, a zsírsavakat és a glicerint,
- a fehérjék bontástermékeit, az aminosavakat, és
- a nukleotidok különféle származékait
felhasználhatjuk egyrészt saját anyagaink (saját fehérjék, szénhidrátok, lipidek és nukleotidok) felépítésére. Az építőanyagként nem hasznosítottakat pedig tovább bontjuk, egészen addig, amíg újra szén-dioxidot és vizet kapunk végeredményként, valamint, és legfőképpen, energiát. Ez a folyamat a sejtlégzés, más néven biológiai oxidáció.
A légzés során tehát az alábbi bruttó folyamat megy végbe:
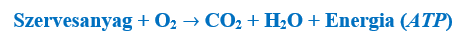
Vegyük észre, hogy a fotoszintézis és a sejtlégzés folyamata éppen egymás megfordítottjai, a különbség csak az irány, valamint az, hogy az „energia” tag eltérő minőségű (mennyiségileg egyébként pontosan megegyeznek). A légzés során tehát azt a szervesanyagot bontjuk le (még a szénvázat is), amelyet valamikor az autotrofok készítettek (noha lehet, hogy egy másik heterotrof állat testével vettük magunkhoz). Ehhez azt az oxigént használjuk fel, amelyet az autotrofok a fotoszintézis során juttattak a környezetbe. (Az oxigénből ugyanúgy víz lesz, mint ahogy előtte volt, és a tápanyag szénvázából is ugyanúgy szén-dioxid lesz, mint ahogy azt az autotrofok felvették.) Az egész sejtlégzési folyamatból számunkra tehát az a fontos, hogy a kémiai kötésekben tárolt – napenergiából származó – energiát felszabadítsuk, valamint felhasználható és tárolható formájúvá alakítsuk.
Erre szolgál az adenozin-trifoszfát, röviden ATP molekula. Az ATP egy olyan egyszerű nukleotid, mely három foszfátcsoportja (PO43-) közti kötéseiben nagy mennyiségű energiát tárol. A foszfátcsoportok egyikének leadásával – reverzibilis módon – adenozin-difoszfáttá (ADP) alakul, és közben, egy kapcsolt reakció számára energiát szolgáltat (1. ábra).
- Ábra: Az ATP–ADP átalakulás sematikus ábrázolása (A foszfátcsoportot az egyszerűség kedvéért P-vel jelöljük, a Pi pedig a szervetlen – inorganikus v. anorganikus – foszfát elnevezésre utal)
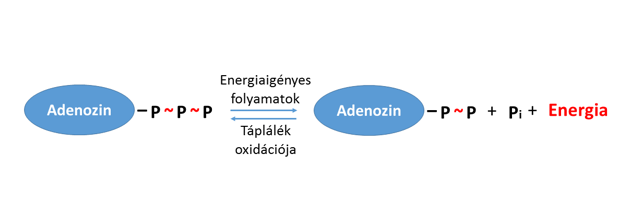
1. Ábra: Az ATP–ADP átalakulás sematikus ábrázolása (A foszfátcsoportot az egyszerűség kedvéért P-vel jelöljük, a Pi pedig a szervetlen – inorganikus v. anorganikus – foszfát elnevezésre utal)
Azt az energiát tehát, ami az ATP → ADP + Pi folyamatban felszabadul, egy másik reakció használja fel. Sejtjeinkben rengetegféle energiaigényes reakció zajlik, de a mérleg másik serpenyőjében mindig az ATP molekula bontását találjuk. Ezt úgy is megfogalmazhatjuk, hogy az ATP az emberi testben egyetemes „fizetőeszköz”, ugyanis minden energiaigényes biokémiai átalakuláshoz ez a molekula biztosítja az energiát. Ez egyébként nemcsak az emberi szervezetet, hanem az egész élővilágot tekintve is érvényes: a baktériumoktól az emlősökig majdnem minden energiaigényes anyagcserefolyamatban ATP-vel fizetnek. A molekula kulcsszerepét és mindenütt jelenlevőségét jól mutatja, hogy szokványos tevékenységeink közben minden nap nagyjából a testtömegünkkel egyező tömegű ATP-t alakítunk át (pl. Törnroth-Horsefield, Neutze 2008).
A sokféle reakció tehát ATP-t használ el, az ATP-fogyasztás ezért az érem egyik oldala. A másik oldalon az ATP regenerációja folyik. Ez főleg a mitokondrium nevű sejtszervecskékben megy végbe. A mitokondrium tartalmazza azokat a molekulákat és reakciókörülményeket, melyek lehetővé teszik, hogy a szervetlen foszfátcsoport (Pi) újra az ADP-re kötődjön.
Izomsejtjeink összehúzódásához is ATP-bontás szükséges: emlékezhetünk, hogy a miozin egy ATP-áz enzim. Ezért aztán nem véletlen, hogy a sporttevékenység alfája és omegája az, hogy adott idő alatt megfelelő mennyiségű ATP-t állítsunk elő. Az alábbiakban áttekintjük, hogy a szervezetünk hogyan termeli meg az ATP-t.
2.2. A szénhidrátok bontása: a sejtlégzés
Az energiatermelés tehát mindig szervesanyag bontásából indul ki, és oxigént igényel (ld. a légzés bruttó egyenlete). Sejtjeink legfontosabb, „üzemanyag” értelmű tápanyaga, a hat szénatomos glükózmolekula a sejtplazmában jelen lévő enzimek segítségével több lépésben két darab, egyenként három szénatomos egységre, piroszőlősavra (melynek vizes oldatban ionja a piruvát) bontódik. A folyamatot glikolízisnek nevezzük. Ennek során 1 molnyi glükóz bontásából 2 molnyi ATP keletkezik.
Glikolízis:
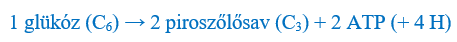
A piroszőlősav aztán a mitokondriumba lép be, és megkezdődik a végső feldarabolódása. Ennek során először egy szénatomot veszít, és acetilcsoporttá (C2) alakul. Ez azért fontos, mert egyébként az összes más típusú szervesanyag (lipidek, fehérjék és nukleotidok) bontása végül a két szénatomos acetilcsoportba torkollik.
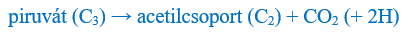
Az acetilcsoport ezután a citromsavciklus nevű körfolyamatba lép be, amelyben egy négy szénatomos vegyülethez kapcsolódik, ami így hat szénatomossá alakul, és miközben több lépésben visszaalakul négy szénatomossá, egy-egy szénatomot elveszít. Tehát a citromsavciklusban végső soron az acetilcsoport szénatomjai közötti kötés felbomlik, és 2 (molnyi) CO2 molekula keletkezik. Ezt a vér a tüdőnkbe szállítja, majd onnan kifújjuk, vagyis visszaadjuk a környezetnek olyan formában, amelyben a növények felvették és fel is vehetik újra.
A folyamat záróakkordja a terminális oxidáció. Ebben a citromsavciklus során és a korábbi reakciókban felszabadult hidrogén elektronokra és protonokra bomlik, miközben az elektronok egy egymás után kapcsolt reakciójú molekulákból álló láncra (elektrontranszportlánc vagy légzési lánc) kerülnek, végül pedig a vérből a sejtekhez diffundáló oxigén veszi fel őket. Az oxigén az elektronokkal együtt a protonokat is felveszi, s így vízzé oxidálódik, ami nagy mennyiségű energia felszabadulásával jár. Lényegében ez az energia használódik fel az ATP újraképzéséhez (ATP-szintetáz enzim: ADP + P → ATP), míg az energia egy része – több mint fele – hővé alakul. Ez a hő energetikai szempontból tekintve veszteség, hideg időben, hőtermelés szempontjából nézve pedig nyereség. (Az élővilágban energiatermelés [ATP-termelés] céljából az oxigén egyetemes elektronfelvevő molekula, azonban léteznek más megoldások is: egyes – pl. mélytengeri hőforrásokban élő – baktériumok szulfátot vagy nitrátot használnak fel anaerob légzésük elektronakceptoraként.)
Végső soron egy molnyi glükóz teljes elégetésével (oxidációjával) összesen 38 mol ATP keletkezik, ami azonban csak elméleti érték, melyet a sejt nem ér el, mert közben az oxidáció bizonyos folyamatai felhasználnak ebből a mennyiségből.
E folyamat ismeretében érthetővé válik, hogy miért fontos oxigénnel ellátni izomsejtjeinket: a mozgáshoz az izmoknak össze kell húzódniuk, ehhez ATP kell, annak a keletkezéséhez pedig oxigénre van szükség. Minél tovább tudjuk tehát biztosítani a kellő mennyiségű oxigént és az oxidáció tápanyagát, annál tovább tudunk ATP-t előállítani. Ha a tevékenység során elegendő oxigén áll rendelkezésre az izomsejtekben, akkor úgynevezett aerob körülmények között használjuk izmainkat.
2.3. Egy alternatív út: az erjedés
A fent leírt bontási folyamatnak a különféle enzimeken és kiindulási anyagokon kívül tehát a legfontosabb feltétele az oxigén jelenléte. Ha nincs a mitokondriumban elég oxigén, akkor nincs, ami a folyamat legvégén felvegye az elektronokat. Így az elektrontranszportlánc részlegesen „bedugul”, és a növekvő ATP-igényt a rendszer nem tudja kielégíteni. Az oxigén relatív hiányában folyó aktivitást nevezzük anaerob tevékenységnek.
A szervezetben erőteljes mozgás közben azonban egy idő után több ATP-re volna szükség, mint amennyit meg tudunk termelni annyi oxigénnel, amennyit képesek vagyunk felvenni. Az energiatermelés azonban szerencsére nem áll le: a glikolízisben előállított piroszőlősav egy része elegendő oxigén hiányában nem a terminális oxidáció felé haladó anyagcsere-útra lép, hanem az erjedésnek nevezett folyamat felé. Az erjedés alacsony hatásfokú energiatermelő folyamat, mert a teljes oxidációhoz viszonyítva ebben körülbelül 1/15-nyi mennyiségű ATP keletkezik.
Az erjedésnek két fő fajtája van az élővilágban attól függően, hogy mi az a termék, amivé a piroszőlősav átalakul. Az élőlények egy részében, mint például az élesztőgombákban a termék etil-alkohol (C2) + CO2. Ezt alkoholos erjedésnek nevezzük. Az emberi vázizomsejtekben a termék a tejsav (C3), vizes oldatban képződő ionját laktátnak nevezzük.
Az erjedés során lényegében a glikolízisben keletkezett piruvát egy olyan termékké konvertálódik (tejsavvá), amely a sejtből könnyen elszállítható, és felveszi a glikolízisben felszabadult H-atomokat. Erre azért van szükség, mert ha valamely reakció terméke túlzottan felszaporodik, akkor ez a reakciót leállítja. Ezért, ha a piruvát átalakul és elszállítódik, nem akadályozza újabb piruvátmolekulák képződését, és a H-atomokat felvevő molekulák is újabb hidrogénatomokért állhatnak sorba a következő glökózmolekula glikolitikus hasításakor.
Az erjedés során glükózmolekulánként mindössze 2 ATP-nyereséggel számolhatunk, azzal, ami a glikolízisben keletkezett.
Az erjedés haszna tehát az, hogy
- az energiatermelés a megnövekedett oxigénigény kielégítő fedezése nélkül is tud folyni, és
- gyorsabban lezajlik, mint a teljes oxidáció.
E tulajdonságai engedik meg azt, hogy az izmok nagy erőkifejtések során a sejtlégzés mellett az erjedést kiegészítő ATP-termelő forrásként használják. Az erjedésnek azonban ára van, ugyanis az előállított tejsav csak ideiglenes megoldást kínál az oxigénszegény időszakra, de energetikai szempontból nézve „adósságot csinál” (ld. még később). Emellett a tejsav túl magas koncentrációban veszélyezteti a szervezet működését, úgyhogy a keletkező tejsavval sejtjeinknek kezdeniük kell valamit. A következő gondolati egység során áttekintjük, hogyan regenerálódik a sejt és a szervezet az erjedés termékeiből.
2.4. A Cori-kör
Az izomsejtekben termelődött tejsavat a máj képes visszaalakítani glükózzá. A teljes folyamat ciklus jellegű: a tejsav az izomból a vérbe pumpálódik, ami a májba szállítja azt, és az itt található enzimek a tejsavból néhány lépésben újra glükózt állítanak elő (glükoneogenezis), ami ezután ismét a vérbe kerül, ahonnan újra fel tudják venni az izmok, melyek aztán újra csak tejsavvá alakíthatják anaerob energiatermelésük során (2. ábra).
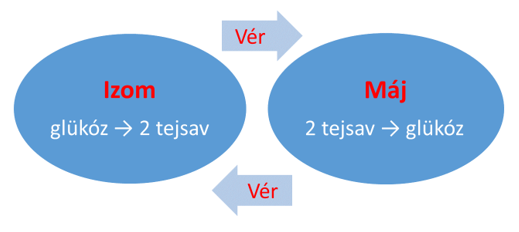
2. Ábra: A Cori-kör
A Cori-kör jelentősége abban áll, hogy
- eltávolítja a túl magas koncentrációt elérő tejsavat az érintett izomsejtekből, és
- újra felhasználható tápanyaggal látja el az izmokat.
Ahogy említettük, energetikai szempontból azonban az erjedés összességében adósságot termel. A máj a glükózzá történő alakítást 6 ATP terhére tudja elvégezni, vagyis a glikolízis 2 ATP nyereségét levonva, a tejsavas erjedés glükózmolekulánként 4 molekula ATP veszteséget produkál, tehát a ciklus nem tud a végtelenségig zajlani. Ezt az ATP-mennyiséget természetesen meg kell később termelni a mitokondriumok terminális oxidációjának segítségével, ami ezért többletoxigén felhasználását igényli a testtől.
A Cori-kör ezért tulajdonképpen az izmok nagy intenzitású működésének az árát ideiglenesen a májra hárítja át, végső soron pedig a teljesítmény csökkentének (vagyis az ATP-igény csökkenésének) időszakában a szervezetnek vissza kell fizetni a kialakult adósságot. A folyamat tehát nem jelent csodaszert, csupán segít túlélni azt a periódust, amíg relatíve túl nagy az energiaigény az adott izmokban.
2.5. A tejsav mint üzemanyag
A Cori-kör mellett a vázizmok is képesek visszaalakítani a tejsavat piroszőlősavvá, és aztán a mitokondriumban teljesen lebontani szén-dioxiddá és vízzé, miközben ATP-t állítanak elő az aerob sejtlégzés során (pl. Bangsbo et al. 1995, Garnier et al. 1996). Ilyen szempontból tehát a tejsav a glikolízisben képződött hidrogénatomok számára csak egy ideiglenes „megőrzőhelyet” biztosít, de a hidrogénatomoknak előbb-utóbb vissza kell térniük „rendeltetési helyükre”, vagyis az energiatermelés aerob útján kell találkozniuk az oxigénnel, vizet képezve (3. ábra). Felmerülhet a kérdés, hogy hogyan végzi el ezt az aerob oxidációt az izom, hiszen amikor tejsavat termel, akkor éppen nem képes elégséges szintű aerob energiatermelésre, mert nincs benne elég oxigén. Természetesen az izom nem is ebben az időszakban égeti el a tejsavat, hanem akkor, amikor a munkaintenzitás csökken – vagy pedig nem azokban az izmokban zajlik le az égetés, amelyek megtermelték a tejsavat, hanem azokban, amelyeknek még van aerob ATP-termelő kapacitása, mert nem dolgoznak nagy intenzitással.
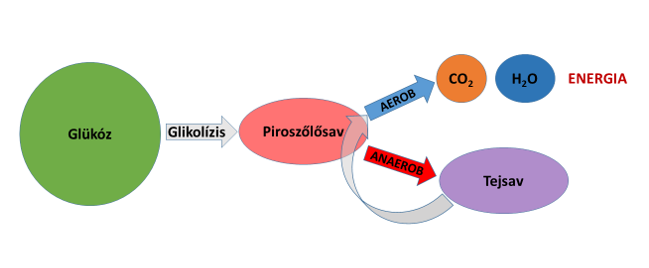
3. ábra: Az izmokban az anaerob erjedésben képződött tejsav előbb-utóbb az aerob energiatermelés útjára lép
2.6. Az izom erőforrásai a tartalékanyagok szempontjából
2.6.1. Az izmok saját energetikai tartaléka: a kreatin-foszfát
Az izomsejtek rendelkeznek egy olyan, közvetlen ATP-tartalékkal is, amely rövid időre pótolja az elhasznált ATP-t. Ez a kreatin-foszfát nevű molekula, amely a keletkező ADP-t regenerálja foszfátcsoportja átadásával:

Így az izomsejtek újabb összehúzódást kezdhetnek meg. Az ezzel párhuzamosan, illetve ezt követően (a glikolízisben és a terminális oxidációban) keletkező ATP segítségével aztán a molekula visszaalakul kreatin-foszfáttá, az előbbi folyamat megfordításaként (4. ábra). Ez a tartalék az izmokban mindössze 10-15 másodperces maximális erejű tevékenység ATP-igényének fedezésére elegendő (pl. Fonyó 2014).
A leadott teljesítmény szempontjából ez a kémiai raktár tehát egy az erjedésnél sokkal gyorsabb, ám sokkal hamarabb ki is merülő, nagyon fontos kiegészítő mechanizmust képvisel.
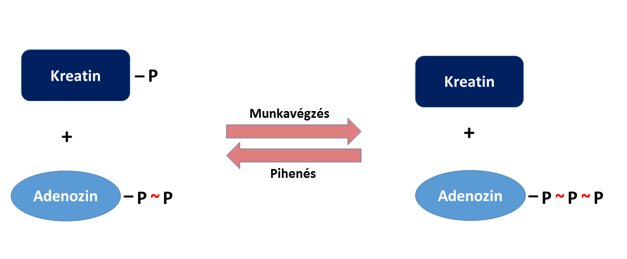
4. Ábra: Egy enzim (kreatin-kináz) a kontrakció alatt ATP-vé alakíthatja az ADP-t a kreatin-foszfát foszfátcsoportjának segítségével, pihenés alatt pedig a többlet-ATP terhére a kreatinból kreatin-foszfátot képez
0.0.1 2.6.2. Az izom tápanyagfelhasználása és saját tápanyagtartalékai
Az izmok tápanyagfelhasználására jellemző, hogy nyugalomban és terhelés alatt is döntően kétféle szervesanyagot oxidálnak: szénhidrátokat és zsírokat (a zsírsavak ugyanúgy acetilcsoportokká bomlanak le, mint a glükózból keletkező piroszőlősav, majd teljesen oxidálódnak szén-dioxiddá, vízzé, és ATP termelődik közben). E tápanyagok felhasználási aránya viszont eltér az edzés intenzitásától és időtartamától függően.
Az izmok zsírsavellátásának javarészét a zsírszövet zsírjának (tömegét tekintve változó, kb. 5–40 kg) bontása fedezi, de maga az izomszövet is rendelkezik egy kisebb (300 g) zsírraktárral (intramuscularis zsír), ami az edzés során elkezd lebomlani (Jeukendrup 2003, Kiens, Hawley 2011). Ennek szerepe leginkább a közepesen intenzív, hosszú ideig tartó (több mint 90 perc) edzés során van, amikor az összes energiafelhasználás negyedét biztosítja. Alacsonyabb vagy magasabb intenzitáson az intramuscularis zsír szerepe csökken az energiaellátásban (Egan, Zierath 2013).
Az izomsejtek emellett jelentős (összesen mintegy 50–900 g) glikogénraktárral is bírnak. A glikogén nevű óriásmolekula egy glükózegységekből felépülő polimer, mely szükség esetén glükózt biztosít az energiatermeléshez (enzimatikus bontás révén). A sporttevékenység során az izom glikogéntartalma is elkezd lebomlani, a keletkező glükóz pedig a teljes oxidáció vagy az erjedés útjára lépve energiát szolgáltat. Az izom glükózszükségletét saját tartalékai mellett a máj glikogénjének (80–100 g) bontásából származó, a vérből felvett glükóz biztosítja. Összességében a zsírraktárak a szervezet energiatartalékának 92-98 %-át, míg a szénhidrátraktárak mindössze 2-8 %-át képviselik (Jeukendrup 2003).
Amennyiben hosszabb ideig (órákon át) tartó munkavégzés történik, az izom saját, helyi tartalékai lassan kimerülnek. Ezután az izom a külső – azaz a vérrel érkező – tápanyagtartalékokra van utalva. Ezek az anyagok a májból származó glükóz (amely a májglikogén elfogytával glükoneogenezissel keletkezik), és a zsírszövet zsírjainak bontásából származó szabad zsírsavak, sőt, az izom a ketontesteket is képes oxidálni. (A ketontestek a zsírsavak lebontásából keletkező acetilcsoportokból jönnek létre a májban, főleg éhezés során, de jelentős fizikai megterheléskor is. Közülük kettőt – β-hidroxi-butirát, acetoacetát – az agy és a szív alternatív tápanyagként képes hasznosítani a glükóz helyett. A harmadik ketontest, az acetoacetátból létrejövő aceton a tüdőn keresztül távozik: „acetonos lehelet”.) E tápanyagok relatív hozzájárulása az energiatermeléshez azonban eltérő lesz: a hosszan tartó sportolás hatására az idő előrehaladtával a szénhidrátok felhasználási mértéke csökken, míg a zsírsavaké növekszik (Jeukendrup 2003).
Amennyiben az edzés intenzitása nő, eleinte mind a szénhidrátok, mind a zsírsavak felhasználási mértéke nő, de egy bizonyos határtól a zsírok felhasználási aránya egyre csökken, míg a szénhidrátoké növekszik (Achten et al. 2002, Jeukendrup 2003, Kiens, Hawley 2011; ld. még később az 5.2 fejezetet). Az intenzitás növekedésével a fő üzemanyag az izomglikogén lesz, ennek felhasználása az intenzitás emelkedésével arányosan növekszik (Egan, Zierath 2013). Megjegyzendő, hogy a szervezet a fehérjéket is fel tudja használni energiaforrásként, ám a fehérjék lebontásából származó aminosav-oxidáció arányaiban nagyon csekély mennyiségű energiát szolgáltat. Edzés alatt az aminosavakból származó energiatermelési hozzájárulás kevesebb mint 1 %-nyi (más szerzők szerint 2–5 % [Moore 2015] és 1–6 % [Tarnopolsky 2004]), és még a kiéheztetett szervezet hosszú időtartamú fizikai igénybevétele alatt is 10 % alatt van (Jeukendrup 2003).
2.7. A sokrétű tejsav
Láttuk, hogy az izommunka során a tejsav fontos szerepet játszik, azonban ezen kívül sok egyéb funkciót is betölt szervezetünkben. A korábban csak egy mellékterméknek tekintett tejsavról az utóbbi években bebizonyosodott, hogy fontos jelátviteli molekula az emberi szervezetben, ami szerepet játszik például a gyulladáscsökkentésben, a sebgyógyulásban, az energiaforgalom szabályozásában, a memória kialakulásában és a daganatok fejlődésében is (Sun et al. 2017). Napi aktivitásunk során a vérbe kerülő tejsav nagyjából negyedrésze az izmokból, negyedrésze az agyból, negyedrésze a bőrből származik, a maradék pedig főleg a vörösvértestekből és a belekből érkezik (Sun et al. 2017).
Fizikai aktivitás folyamán természetesen az izmok tejsavkibocsátása nagymértékben megnő, amit a máj – akár tízszeresére (Sun et al. 2017) is – növekvő tejsavfelvétele kompenzál, hiszen ez a szerv a glükoneogenezisen keresztül képes csökkenteni a laktát túl magas koncentrációját (kisebb mértékben a vese is gyárt glükózt a laktátból).
Ahogy láthattuk, hasonló módon viselkednek azok a vázizmok is, melyek éppen nem dolgoznak magas intenzitással, vagyis aerob energiatermelési tartományban vannak: ők is felveszik a társaik által termelt tejsavat, és visszaalakítják piroszőlősavvá, majd eloxidálják a mitokondriumokban (pl. Bangsbo et al. 1995, Garnier et al. 1996). Ezzel mintegy átvesznek az anaerob zónában tevékenykedő izmok ATP-ben kifejezett adósságából. Mindezek mellett a szívizomzat is felveszi a tejsavat és tápanyagként hasznosítja (ugyanúgy, mint a vázizmok). Ez jelentős energiaforrás a szív számára, hiszen tápanyagellátását nyugalomban kb. 10-15 %-ban, míg fizikai tevékenység alatt kb. 30 %-ban a tejsav fedezi (Sun et al. 2017).
2.8. Az elméleti alapok összegzése a sport szempontjából
A fentiek fényében most már összefoglalhatjuk, hogy a fizikai munkavégzést – és így a sporttevékenységet – alapvetően milyen tényezők határozzák meg az anyagcsere oldaláról. Ha a tevékenység nem túl intenzív, vagyis elegendő oxigén áll rendelkezésre, akkor az energiatermelés legfőképpen a glükóz és a zsírsavak oxidációjával fedeződik (glükózmolekulánként névlegesen 38 ATP molekula keletkezését eredményezve). Ez a folyamat eleinte az izom glikogéntartalmát és szerény zsírraktárát emészti fel, majd hosszabb távon a zsírszövet zsírjának égetésével folytatódik. Az elegendő oxigén pontosabban azt jelenti, hogy oxigénfelvevő képességünk a munka intenzitásához képest kielégítő, azaz „győzzük levegővel” az erőkifejtést. A sportolás során ezt a képességünket jelentősen fejleszthetjük (ld. később). A sportmunka ilyen feltételek között zajlik javarészt aerob tartományban (5. ábra). Ebben a zónában tehát két sarokpont az időtényező és a tápanyagok mennyisége: meddig vagyunk képesek üzemanyagot adni az izmoknak; meddig tudjuk a glikogént, meddig a zsírt biztosítani, és a verseny mely szakaszában muszáj például nagy tápanyagtartalmú italokkal pótolni a kimerülő tartalékokat. Ez tipikusan például a maratonfutók, a triatlonisták, a hosszú távot úszók, az országúti kerékpárosok, az ironman-versenyek résztvevőinek teljesítményi zónája.
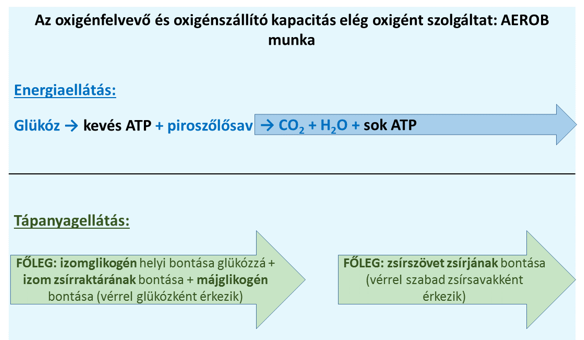
5. ábra: Az aerob energia- és tápanyagellátás
Ha azonban a tevékenység intenzívebb ennél, akkor több oxigénre volna szükség a sejtlégzésben, mint amennyit a vér oda tud szállítani és a sejtek fel tudnak venni. Tehát az aerob kapacitás már nem képes biztosítani a szükséges ATP-termelést, és az energiatermelés egy másik része egyre növekvő mértékben anaerob laktacid, azaz anaerob–tejsavtermelő üzemmódba kell hogy kapcsoljon. Nagyon fontos kiemelni, hogy a sejtlégzés természetesen nem áll le ezalatt, csak a többlet-ATP előállítására kell a sejteknek megoldást találniuk (6. ábra). Az első néhány másodpercre a kreatin-foszfát kellő mennyiségű ATP-t biztosíthat, azonban – a sprintszámok kivételével – a legtöbb sportban ennél jóval tovább van szükség a többlet-ATP-re. Ekkor a glikolízisben keletkezett piroszőlősav nem a mitokondriumokba lép, hanem tejsavvá alakul (vagyis az erjedés indul el), amely kilép a sejtből, és a májba szállítódik, hogy a glükoneogenezisben visszaalakuljon glükózzá (Cori-kör). Egy másik része a szívbe kerül, a többit pedig az éppen nem olyan nagy intenzitással dolgozó vázizmok veszik fel. Amíg a vér által megvalósított laktát–glükóz csere a máj és az izmok között, valamint a szív és a többi vázizom által biztosított tejsavfelszívás és elégetés elégségesen ellensúlyozza a laktátprodukciót, ez a munkaintenzitás fenntartható. A teljesítmény további fokozásával az anaerob energiatermelés generálta tejsav koncentrációja egyre növekszik a vérben. A továbbiakban a teljesítményleadást az határozza meg, hogy milyen magas vér-laktátszintet tolerál a szervezet. Ez a képesség – a laktát-tolerancia mértéke – is fejleszthető.
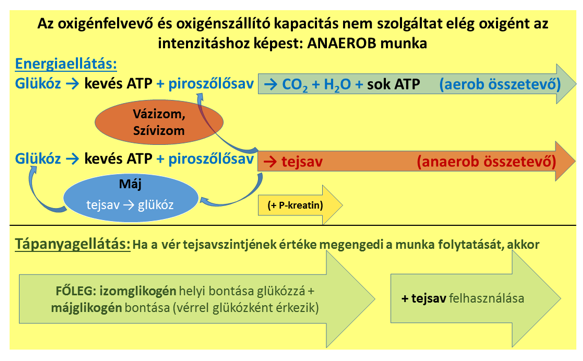
6. Ábra: Az izommunka anaerob teljesítményi tartományai az oxigén- és a tápanyagellátottság szempontjából. Az anaerob munkavégzést bármilyen tápanyag-felhasználási fázisban leállíthatja a vér túl magas tejsavszintje. A kisebb sárga nyíl azt fejezi ki, hogy az ATP-tartalékként szolgáló kreatin-foszfát rövid, korlátozott időszakra biztosíthat energiát a tevékenység nagy erőkifejtéssel végzett fázisaiban. A tápanyagfelhasználásra jellemző, hogy nagy intenzitásnál alapvetően szénhidrátot használ a szervezet, emellett pedig a termelődő tejsavat is igyekszik hasznosítani.
A rendszer harmadik fontos összetevője a foszfokreatin-rendszer. Ez gyors kémiai lépéseken keresztül, nagyon hamar szolgáltat ATP-t a megnövekvő teljesítményigény kielégítésére, de csak nagyon rövid ideig. Ezt a rendszert nevezzük anaerob alaktacid, vagyis anaerob, tejsavat nem termelő rendszernek. Az anaerob alaktacid energiatermelést egyfajta „turbo-fokozatként” képzelhetjük el, ugyanis a folyamatosan zajló aerob ATP-termelés és a gyors tejsavszintézis által produkált ATP-előállítás mellett olyan plusz tartalékot juttat a rendszerbe, amely lehetővé teszi, hogy pár másodpercre ugyan, de jelentősen fokozzuk izomzatunk teljesítményét. Ha ez a tartalék kimerül, akkor pedig a terminális oxidációban keletkező nagymennyiségű ATP-nek vissza kell töltenie a készletet. Ezért a sportolás folyamán ezt a plusz energiaforrást többször is bekapcsolhatjuk, ha figyelünk rá, hogy legyenek gazdaságosabb teljesítményperiódusok is a tevékenység során, melyek alatt visszatöltjük ATP-készletét.
Sok sportélettannal foglalkozó könyvben és magazinban az aerob, az anaerob laktacid és az anaerob alaktacid energiatermelési tartományokat lineáris, szekvenciális egységekként írják le (7. a ábra). E szövegek sokszor arra engednek következtetni, hogy a sporttevékenység során először az aerob rendszer működik, azután pedig az anaerob laktacid rendszer. A foszfokreatin-rendszert pedig gyakran úgy említik, mint a rögtön robbanásszerű teljesítmény leadásával kezdődő aktivitás első szakaszának energiaellátását biztosító, 10-15 másodperces időtartamra szóló tartalékot. Teszik mindezt annak ellenére, hogy évtizedek óta leírták már, hogy a vázizmokban már akkor is működik mind a laktacid, mind az alaktacid anaerob rendszer, amikor az aerob út még bőven képes energiát termelni (pl. Sahlin et al. 1987). Azt szintén hibásan sugallják, hogy nagyobb erőkifejtésnél az anaerob rendszer átveszi az aerobtól az energiatermelést (7. b ábra) – ebben az esetben az erőteljesen dolgozó izmoknak nem volna szükségük oxigénre, csak miután véget ért az intenzív igénybevétel. A kreatin-foszfát tartalék sem csak a terhelés végeztével, a pihenés során tölthető fel, ugyanis az általa szolgáltatott plusz energia nem csak egyszer vethető be az aktivitás során. Ezek az elgondolások ellentétesek a tapasztalatokkal, és logikátlanok is.
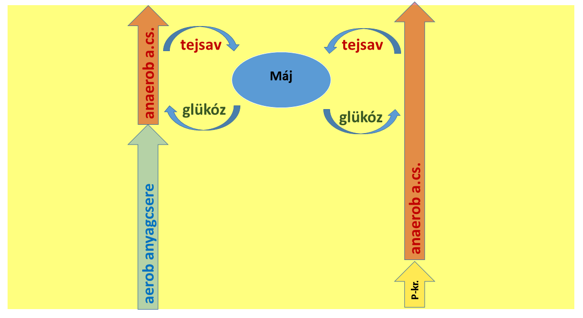
7. a Ábra: Az energiatermelő útvonalak HIBÁS modellje. Bal oldalon az aerob –tejsavtermelő, jobb oldalon a kreatin-foszfát – tejsavtermelő feltételezett működési szekvenciák folyamatvázlata látható
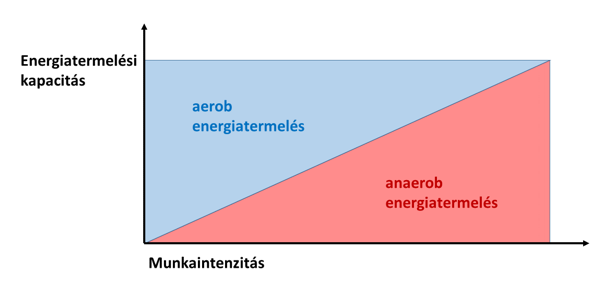
7. b ábra: A HIBÁS modell szerint az anaerob energiatermelés fokozatosan átveszi az aerob anyagcsereút szerepét az intenzitás növekedésével
Az eddigiekben áttekintett ismeretek alapján logikusan egy sokkal életszerűbb rendszer körvonalai bontakoznak ki (lásd például Jamieson 2017). A három energiatermelő rendszer (aerob, anaerob laktacid és anaerob alaktacid) mindvégig egymás mellett működő anyagcsereutak, ám egymáshoz viszonyított jelentőségük eltér. Az energiatermelés alapját az aerob rendszer szolgáltatja. Nemcsak azért van ez így, mert az aerob rendszernek nagy és gazdaságos az ATP-előállító képessége, hanem mert a munkaintenzitás fokozódásával egyre erőteljesebben termelődő tejsav által jelentett energiaköltséget is ő tudja csak visszafizetni. Az anaerob laktacid rendszer előállította laktát vagy a Cori-körben, vagy a szívizomban, vagy pedig a vázizmokban hasznosul, de mindenütt aerob folyamatok során (a máj glükoneogenezisének ATP-költsége, szívizomban vagy vázizomban folyó oxidáció).
Tehát a teljesítmény fokozhatóságának egyik korlátját éppen az jelenti, hogy a sokrétűen ható aerob folyamatok meddig képesek fedezni az anaerob tejsavtermelő út gyors ATP-szintézisének járulékos költségeit. Addig ugyanis szervezetünkben nem halmozódik fel a laktát veszélyes mértékben. Ha a munkaintenzitással efölé megyünk, és a következő, kevésbé erőteljes munkaszakaszban nem tudjuk helyreállítani az adósság okozta kárt (vagy nincs is kevésbé erőteljes szakasz), akkor a felgyülemlő tejsav egyszercsak megálljt parancsol a tevékenységnek. Ha azonban „pihenősebb” szakaszokat teszünk az edzésbe/versenybe, akkor ezek alatt regenerálni tudjuk az anaerob laktacid rendszer előállította tejsav-adósságokat. Amikor szélsőséges mértékben kell fokozni a teljesítményt, akkor az anaerob alaktacid (vagyis a kreatin-foszfát biztosította) rendszerünk „tesz rá még egy lapáttal” az erőkifejtésre, és természetesen ezt a rendszert is visszatölthetjük az aerob rendszer segítségével, egy csökkentebb intenzitású szakaszban. A változó intenzitással dolgozó sportágakban – pl. labdajátékokban és küzdősportokban – ezek alapvető és gyakori módszerek a teljesítményfokozásra; a lineáris, régi modellben mindez nem kap kielégítő magyarázatot. A leírt összefüggéseket szemlélteti a 8. ábra.
8. Ábra: Az energiatermelő rendszerek közötti kapcsolatok
Ami rendszerszinten zajlik, az tehát nem más, mint hogy a nagy kapacitású és sokoldalúan edzhető aerob anyagcsereút mint energiatermelési alap rendelkezik két kiegészítő, de folyamatosan adósságot csináló energiaforrással: a hosszabb távon működő anaerob laktacid és a rövid távon működő anaerob alaktacid rendszerrel. Mindkét plusz energiatermelő ág jelentősen megnöveli az izmok által leadott teljesítményt, ám ezt az aerob rendszer ATP-termelésének a terhére teszik. Az aerob és a két anaerob rendszer közötti viszonyt úgy is leírhatnánk, mintha az aerob rendszer egy autó belső égésű motorja volna, míg az anaerob egységek két teljesítménynövelő plusz akkumulátor. Amikor ezeket igénybe vesszük, veszítenek töltöttségükből, és egy idő után le is merülnek – a feltöltésüket pedig a belső égésű motor tudja elvégezni. Az aerob rendszer tehát folyamatosan dolgozik azon, hogy a pillanatnyi igények miatt működésbe lépő kiegészítő akkumulátorai által létrehozott adósságot visszafizesse (9. ábra). Ha egyikük – a laktacid rendszer – olyan mértékű adósságot csinált, melyet a test nem tud visszafizetni, akkor a tevékenység leáll, vagy pedig katasztrofális következményekkel jár a szervezetre nézve (laktátacidózis; ld. később).
A kiegészítő energiatermelő rendszerek működésére jó példát szolgáltatnak a küzdősportok meneteiben vagy a labdajátékok során jelentkező erőteljesebb szakaszok, amikor a versenyzők „megpörgetik” a mozdulatokat – ilyenkor az aerob motor mellett bekapcsolnak az anaerob akkumulátorok is, de még a komolyabb fáradás megindulása előtt utóbbiak leállnak, és tartalékaik visszatöltődnek az aerob ATP-termelés segítségével. Egy másik példa az aerob-anaerob rendszerek szinergizmusára a sprintfutás, ahol a versenyzők számára az energetikailag gazdaságos erőbeosztás nem szempont, hanem a teljesítmény egy, hirtelen összpontosított erőkifejtéssel érhető el, és utána van idő és mód regenerálni a sejttartalékokat.
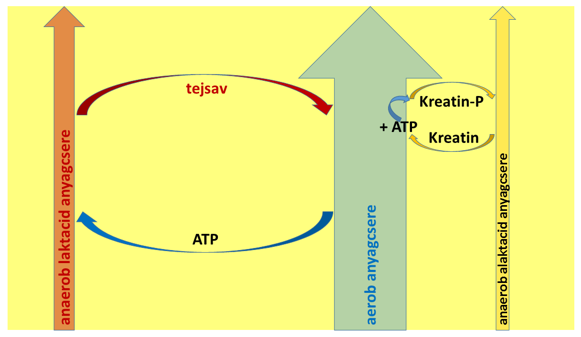
9. Ábra: Az aerob, az anaerob laktacid és az anaerob alaktacid energiatermelő rendszerek közötti rendszerszintű, lényegi kapcsolatok