3. A vázizomrostok élettani tulajdonságai
3.1. A vázizomrostok típusai anyagcsere és működés szempontjából
L-A. Ranvier francia anatómus – aki az idegsejtek axonjait körülvevő velőshüvely befűződéseit is felfedezte – 1873-ban leírta, hogy a nyúl egyes izmai vörösebbek, más izmai sápadtabb színűek (Zierath, Hawley 2004), és ezt a jelenséget később más állatokban is megfigyelték. Ez a jellemzés alkotta az alapját a vörös–fehér izomrost tipizálásnak is. A későbbiekben ez a felosztás finomodott.
A lassú összehúzódású izomrostokban (ezek azonosíthatóak a vörös rostokkal) sok mitokondrium található, ezért nagy teljesítményű oxidatív energiatermelésre képesek, vagyis munkájukat főleg aerob tartományban végzik. Aránylag sok bennük a mioglobin nevű oxigéntároló fehérje, mely a hemoglobin szerkezeti rokona, és átmeneti oxigénhiányos állapotokban kisegíti a sejtet oxigénnel. Glikogéntartalmuk kicsi, ezért a vér tápanyagtartalmára vannak utalva – a lassú izomrostok erezettsége gazdag. Ezek a sejtek egyébként a magas koncentrációjú mioglobin miatt kapták a vörös rostok nevet. Legtalálóbb elnevezésük a lassú-aerob rostok lenne, de a könnyebb megjegyezhetőség miatt hívhatjuk őket vörös-lassú rostoknak is.
A gyors összehúzódású izomrostok egyik fajtája, a gyors–anaerob sejttípus kevés mitokondriumot és mioglobint tartalmaz, de sok bennük a glikogén. Ezek a rostok alapvetően a glikolízisből nyerik az ATP-t, tehát főleg anaerob tartományban dolgoznak (de természetesen aerob oxidációt is végeznek). Ezeket a rostokat írták le fehér izomrostokként, ezt az elnevezést követhetjük mi is.
A harmadik típus az előbbi kettő keverékének is tekinthető, mert az említett molekulákat és sejtalkotókat köztes mennyiségben tartalmazza, összehúzódása gyors, de anyagcseréje inkább oxidatív. Ezek a gyors–aerob rostok. A könnyebbség kedvéért nevezhetjük őket vörös-gyors rostoknak is, de ne felejtsük el, hogy a klasszikusan vörös (tehát vörös-lassú, vagyis lassú-aerob) és a fehér rostokhoz képest ezek a főbb sejtalkotó-jellemzőiket tekintve köztes értéket képviselnek. A három izomrosttípus közötti alapvető különbségeket az 1. táblázat foglalja össze (Herbison et al. 1982; Zierath, Hawley 2004, Egan, Zierath 2013).
(Az izomrostok egy másik szokásos elnevezés szerint I., valamint II.A és II.X [régebben: II.B] típusokba sorolhatóak. Az I. típusú a lassú összehúzódású, a II.A a gyors, oxidatív, a II.X pedig a gyors, glikolitikus rostokat jelöli. Előbbi elnevezéseinkkel élve az I. típus a lassú-aerob [vörös-lassú], a II.A típus gyors-aerob [vörös-gyors], a II.X pedig a gyors-anaerob [fehér] rostoknak feleltethető meg. Ennél finomabb felbontás is tehető különféle molekuláris jellemzők alapján [I., I.C, II.A, II.B, II.C, II.AB, II.AC; pl. Scott et al. 2001], számunkra azonban ezek alkalmazása számottevő haszonnal vélhetően nem járna.)
Kiemelhetjük, hogy a lassú, főleg aerob tartományban munkálkodó rostok abban is fontos szerepet játszanak, hogy a gyors, glikolitikus rostok által termelt tejsavat felvegyék és eloxidálva hasznosítsák, s ezáltal jelentősen hozzájárulnak ahhoz, hogy az aerob rendszer újratölthesse az anaerob rendszer merülő tartalékait.
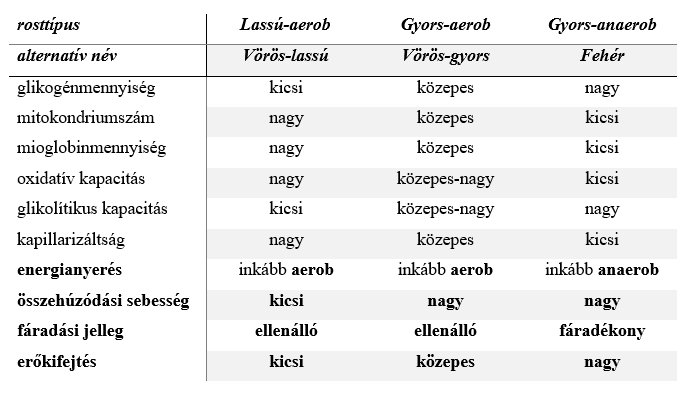
1. Táblázat: A lassú és a gyors összehúzódású vázizomrostok közötti alapvető működési és anyagcserekülönbségek (Herbison et al. 1982; Zierath, Hawley 2004; Egan, Zierath 2013)
3.2. Az izomrostok munka közben
Egy mozgató idegsejt és az általa beidegzett izomrostok együttese alkotja a motoros egységet. Egy-egy motoros egység egyféle működéssel jellemezhető, tehát nagyon hasonló rostokból tevődik össze (Scott et al. 2001). Összehúzódási sebességük alapján a motoros egységek között megtaláljuk a lassú egységeket, a gyors–kitartó, a gyors–köztes fáradású és a gyors–fáradékony egységeket (Scott et al. 2001). A patkányokban, egerekben, tengerimalacokban, csirkékben, stb. leírt, egyértelműen vörös és fehér típusokba besorolható izmoktól eltérően azonban az emberi izmok nem egyféle motoros egységből épülnek fel, hanem izmaink ilyen szempontból kevertek (Johnson et al. 1973; Herbison et al. 1982, Zierath, Hawley 2004): mindig tartalmaznak gyors és lassú egységeket is, de eltérő arányban. Főként lassú rostokat találunk a kitartó működést igénylő izmokban, mint például a testtartásért felelős antigravitációs izmokban; ezzel szemben gyors rosttúlsúly figyelhető meg gyorsan mozgó végtagizmainkban (Johnson et al. 1973).
Alapvető különbség még a lassú és a gyors rostok alkotta motoros egységek között az is, hogy a lassú egységek általában kisebbek, mint a gyorsak. Biológiai tanulmányainkból emlékezhetünk, hogy a mozgások során először a kisebb motoros egységek lépnek működésbe (ugyanis ezek motoneuronjainak az ingerküszöbe alacsonyabb), s csak később az egyre nagyobb egységek (a motoros egységek méret szerinti belépésének elve). Ez azt jelenti, hogy a lassú rostok motoros egységei általában hamarabb kapcsolódnak be a kontrakcióba, és csak a nagy erőkifejtéssel járó, gyors mozdulatoknál lépnek be a nagyobb, gyors rostokat tartalmazó egységek. Egy érdekesség, hogy ezért két, izomtömeget csökkentő hatású folyamat eltérőképpen hat a gyors és a lassú rostokra. Az immobilizáció – pl. gipszelés – hatására elsősorban a lassú rostok szenvednek leépülést. Ennek az lehet a magyarázata, hogy a gyors egységek eleve „megszokták”, hogy csak ritkábban ingereljék őket, lévén életük nagy részében inaktívak, így a stimuláció hiánya inkább a lassú rostokat érinti, amelyek szinte folyamatos ingerlést kapnak a mindennapok során. A másik hatás, az öregedés viszont éppen a gyors rostok mennyiségében okoz nagyobb arányú csökkenést, ami magyarázhatja a gyorsaság és az erő magasabb fokú csökkenését az időseknél (Radák 2016). Ezért kiemelten fontos az időskori fizikai aktivitás, sőt, az erőedzés is.
Az egyes izmokat jellemző rosttípus-eltérések egyének között is megfigyelhetőek, hiszen ugyanazon izomban is más és más a rosttípusok aránya a populáción belül (pl. Zierath, Hawley 2004). Ez a magyarázata annak, hogy egyesek miért inkább állóképességi sportokban, mások miért inkább sprintszámokban mutatnak tehetséget. Ugyanakkor az is elmondható, hogy kitartó edzésmunkával egy bizonyos mértékig saját izmainkban is megváltoztathatóak a rostarányok (pl. Scott et al. 2001; Handschin, Spiegelman 2008; Kelly 2012; Viollet 2018) – például hosszútávfutók lábizmaiban a sportolás hatására a vörös izomrostok aránya az évek alatt nő, míg a sprinterekében csökken. Különféle sportágakban tevékenykedő sportolók izomösszetételének vizsgálatai nagyjából 40 évvel ezelőtt megmutatták, hogy az állóképességi és a rövid távokban teljesítő versenyzők izmainak rostösszetétele eltér egymástól (pl. Costill et al. 1976), megerősítve azt a korábbi nézetet, hogy a sikeresség egyik fontos elemét képezik az izmaink genetikai adottságai; jóllehet, a sportolás hatására végbemenő változásokat ezek a vizsgálatok nem vették figyelembe. Fink és szerzőtársai (Fink et al. 1977) elit (férfi) hosszútávfutók, jó, de nem világklasszis hosszútávfutók és edzetlen férfiak lábikraizmainak összetételét hasonlították össze, és azt találták, hogy a csúcs-atléták izmaiban jóval nagyobb volt a lassú rostok aránya (79% ± 3,5%) a jó futókéhoz (61,8% ± 2,9%) és az edzetlenekéhez képest (57,7% ± 2,5%) is. Azonban közelebbről szemügyre véve az adatokat kiderült, hogy az egyének között megfigyelhető teljesítményi eltéréseket csak a rosttípusok eltérő arányával megmagyarázni nem lehet. Például az elit sportolók között ha összehasonlítjuk a két nagyon hasonló legjobb maraton-eredménnyel rendelkező futó adatait, akkor azt látjuk, hogy lassú rostjaik arányát tekintve óriási a különbség közöttük: 98% és 50% (Fink et al. 1977). Ezért levonhatjuk a következtetést, hogy a rosttípusok aránya nem feltétlenül jelent biztos alapot a teljesítmény előrejelzésére, hanem egyéb jellemzőket is fontos figyelembe venni (Zierath, Hawley 2004).
Nem mindegy például, hogy a versenyző mennyi levegőt képes egyszerre beszívni, milyen pulzustérfogatot tud teljesíteni a szíve, milyen szállítókapacitású érhálózat juttatja el az oxigént a szövetekhez, mennyi oxigént képes az izomzat az érkező vérből kinyerni, és – nem utolsó sorban – milyen a szervezete tejsav-tűrőképessége. A továbbiakban áttekintjük ezeket a limitáló tényezőket és összefüggéseiket.