2. The functioning and adaptability of the locomotor system
We strive to maintain the various parameters of our organism in different domains, so ensuring our survival. If we go beyond the usual parameter ranges, an adaptation response begins. If the displacement is too fast and/or powerful and is not followed by a rapid recovery, the individual will be in a state of shock and, without help, may lose his or her life. However, if we move out of the optimum ranges gradually, in small steps, and we exercise in this way regularly and enough times, we may be surprised at what extreme conditions we will be able to tolerate. We will be able to maintain our attention for a long time, to transform our sleep homeostasis, to endure extreme temperature values without damage, to tolerate water-deficient conditions, and to withstand or recover from extreme, damaging physical effects, and so on. Of course, this general adaptability also extends to our locomotor system. This is one of the most important foundations of an outstanding athletic performance, besides the mental and cardiovascular adaptation. The latter will be discussed in the physiological chapters in the next semester. The present subject aims to examine muscles, the neuromuscular coordination and joints, to this end.
2.1. Muscles and tendons
2.1.1. Basics of muscle function
Muscle is the main structure for locomotion of the animal-type life form on earth. The smallest muscle in the animal world is less than one millimetre long (microscopic crustaceans), while the longest ones are in the meter range (whales’ muscles). This is roughly a hundred thousand-fold difference in length, and about a quadrillion-fold (million billion, i.e. 1015) in volume (Vogel 2013).
Human muscle tissues can be divided into three groups, as we can remember: the smooth muscles providing visceral motility, the heart muscle, and the skeletal muscle tissue that carries out voluntary movements. Of course, in sports we mainly use the latter, because our skeletal muscles are formed from striated muscle tissue. Muscle contraction is made possible by the fact that the protein molecules of a specific structure, located in the muscle cell in bundles, the actin and the myosin, slide past each other (Figure 2). The actin fibres are fixed to a sheet-like, transverse protein structure, the so-called Z-disc (or Z-line), so, by moving they move these anchoring surfaces together. Ultimately, since the Z-discs are also fixed to the cell membrane, the actin-myosin protein filaments, by their sliding, contract the muscle cell. This process takes place at the expense of ATP, since myosin acts as an ATPase enzyme, that is, it is capable of converting ATP to ADP, while undergoing spatial transformation that results in actin filaments sliding between the myosin filaments. Thus, the release of chemical energy is transformed into mechanical energy. Replacing the ATP supply and the ATP reserve is a key issue for long-term muscle usage. We will deal with this in more detail in the physiology section.
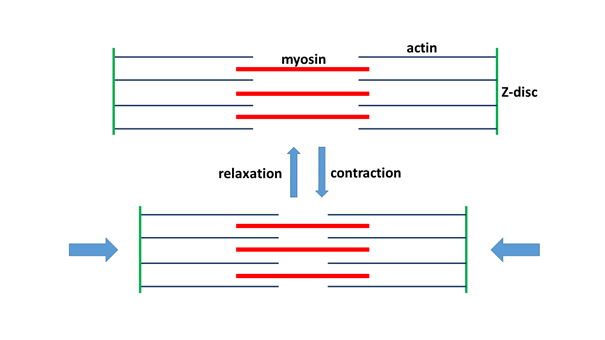
Figure 2. The contraction of the muscle cell is ensured by the sliding together of actin and myosin(Figure: Author’s own production.)
The skeletal muscle fibres are formed by the fusion of several cells, so they contain several nuclei, which are located on the periphery of the cells, similar to the large number of mitochondria that produce energy (in the form of ATP). Almost the entire interior of the cell is filled with bundles of actin-myosin molecules.
Fibres of the skeletal muscles form three main types, which have different optimal power ranges due to their different energetic characteristics. Each of these fibre types can be found in each of our skeletal muscles (one type per motor unit), but their proportions are different (they are discussed in more detail in Sports Physiology).
2.1.2. Biomechanical peculiarities of muscle tissue
During the discussion in Section 2.1.2, we follow the thread of Steven Vogel’s book on biomechanics (Vogel 2013).
2.1.2.1. Contraction
Although we usually talk about contraction when dealing with muscle work, the muscle is never literally contracted. The actin and myosin filaments do not contract, but when examined at the macroscopic level, the muscle does not contract either, i.e. it does not diminish. The muscle is actually shortened while it swells – that is, it undergoes a change which is isovolumetric (i.e. without a change in volume). Imagine – as a simple proof on the whole organism level –, that we take exactly as much air into our lungs to enable us just to remain on the surface in a pool of water. We suddenly pull together as much muscle as we can. What would we expect if the muscles really “contracted”? Our volume would decrease, so our density would increase, so we would sink. However, this does not happen, because nothing happens: we continue to float on the water – so the muscles do not really “contract”.
The role of the muscles could be more appropriately formulated as pulling the two end structures towards each other while swelling. However, this cumbersome wording would hamper communication, so we can still use the term “contraction”.
The basis of the shortening is therefore the actin fibre displacement due to the ATP cleavage by myosin molecules. This molecular force is very small, but with many of these enzymes and enzymatic reactions placed side by side, the force multiplies. Thus, the muscle cell is actually an enzyme bundle well-organized in terms of space and function. However, since they are molecules that move on each other in an ordered manner, it is also necessary for the organized operation that they can move only in the desired direction during movement, and return precisely to their starting position. This is facilitated by a giant protein molecule that, coupled to the Z-plates (each anchoring on the opposite Z-disc, and connected to each other on the other end), flexibly holds the strictly ordered filaments in place. This protein is titin; and it will be discussed later, together with the elastic elements.
The skeletal muscles are capable of pulling with about a force of 200 kilopascals due to this precise protein assembly. In stating this, it is not the absolute figure which is significant, but the fact that the tensile strength of the joints’ collagen is about 500 times that value (Vogel 2013). This explains why the bulky, swollen muscles can connect to the bones with thin tendons.
2.1.2.2. Tendons
The muscles are thus connected to the bones by the tendons. The tendon is a structure formed by the so-called dense connective tissue in which collagen fibres of great flexibility and tensile strength run in parallel. Collagen is a protein molecule that consists of fibres running in a triple helix structure, and is a universal connective tissue component in the human (and generally in the mammal) body. We can find it in the skin, muscles, bones, cartilage, tendons, and ligaments, but also in the cornea of the eye, in the walls of the blood vessels, or in the dentine substance of the teeth.
High tensile forces induce solid bodies to produce a generally characteristic reaction. They extend to a certain value of force and then, with the cessation of the force, regain their former shape and volume, i.e. they are flexible (the tension of the material increases, and its elongation increases; in this elastic phase there is a proportionate and disproportionate tension phase, but for the sake of simplicity we will not deal with this in detail). With greater force, the material just flows, becomes plastic (its tension decreases, but its elongation continues), and with even greater force it tears when the tensile strength value is reached. The connective tissues work in a similar manner, but they do not exert any flexible force as a result of a small force, but simply stretch; then the flow phase follows, and finally the tear (Katics, Lőrinczy 2010).
The tensile strength of the tendons is so great that when too great a tensile force acts on the muscle, it tends to break the muscle, or the tendon tears a piece out of the bone; but the tendon itself holds on (the tendons are connected to the bone through a cartilaginous cellular transition and their collagen fibres penetrate into the bone tissue, which provides very strong adhesion to the tendons; the muscle-tendon transition is also a similarly strong structure).
2.1.2.3. Muscle and power
It is no accident that the muscle is such a widely used structure in the animal world: in terms of output, the muscle is the absolute biological champion among multicellular organisms with its maximum average value of 200 watts/kilogram (the second, the eukaryotic cilium is far behind, at about 30 watts/kg; Vogel 2013). However, this value is far from its real performance; neither the muscle, and even less a whole human being, can produce this much output, but only a fraction – in fact, less than a tenth. Therefore, it can be said that the power delivered is not limited by the capabilities of actomyosin bundles, but by other features of the system operation, such as the nutrient and oxygen supply of the muscles, and the constraints of heat loss.
Of course, muscle performance also depends heavily on the length of time it needs to work. It can provide a lot of power for a while, but if it has to be maintained for a long time, its performance will be drastically reduced. It is interesting to note that, according to Wilkie’s calculations (Wilkie 1960), a trained athlete can deliver an output of roughly 4500 watts for about one second. If s/he needs to keep working for five seconds, then the power drops to only 1500 watts, to 370 watts for five minutes, and to 370-300 watts for a few hours, and if s/he has to work all day, the power is just around a maximum of 150 watts, which is about 2 watts/kilogram (the listed values are for a variety of tasks, not for the same task). For the sake of completeness, it should be noted that since the above numbers refer to trained athletes, people with average abilities can produce less than 70-80% of the above performance values.
Today, in racing sports, especially for road-race cyclists, the so-called Wattmeter is used in sports competitions. This is a tool to indicate to an athlete how to schedule his/her effort at each attack or reaction, without jeopardizing his/her race by exerting too much energy.
Compared to mammals, humans perform quite well in long-term work, producing about 2 watts/kg (Vogel 2013). But when we compare ourselves to flying animals, we are very far behind. According to the results obtained from the calculations of the oxygen consumption of birds, a 10 dkg bird delivers about 24 watts/kg in flight (Butler 1991), which is a multiple of a human performance. But if we look at arthropods, we can go even further on the imaginary scale: a bumble bee produces an output of about 100 W/kg in flight (Ellington et al. 1990). Swimming seems to be much less expensive than flying: for example, red salmon work at about 0.165 W/kg during economic swimming (Brett, Glass 1973; Vogel 2013), which is much less than the approximately 2 W/kg of human power output.
2.1.2.4. Negative work: eccentric muscle contractions
When a muscle is pulled by a greater force than it can exert, it is in tension, but at the same time it lengthens and absorbs mechanical energy. This work during lengthening is called eccentric contraction, as opposed to concentric contraction with muscle shortening. (The term ‘eccentric’ implies that the ends of the muscle move away from its centre, while ‘concentric’ expresses a movement towards the centre.) If we consider this in more detail, a remarkable feature of muscles’ work is that they can also exert a force while lengthening. This operation is extremely important because, although we tend to think of muscles as “positive” workers, i.e. agents that move objects, the muscles, with lengthening work, can act as brakes on the one hand, and on the other hand, they also store partially recoverable flexible potential energy (Lindstedt et al. 2001). These are the essential elements of harmonious movement.
This kind of work, which is actively “contracted” during prolongation, is particularly effective in muscle building (Lindstedt et al. 2001, Roig et al. 2009), as it strongly stimulates the synthesis of new muscle tissue: weight lowering is more strengthening than weightlifting. It is important to note, however, that this is not about lowering an object raised by positive work, but about an object that we would not otherwise be able to lift. Thus, we urge the muscles to do a job that exceeds their strength, and can only exert force when working with eccentric contraction. For example, an excellent leg muscle builder is a bike whose pedals are rotated backwards continuously by a motor, and the athlete continually tries to roll them forward, in vain. This machine makes the legs of the cyclist do permanently negative work (Lindstedt et al. 2001, Vogel 2013). With such a machine, Lindstedt and his colleagues trained their subjects for eight weeks, and the strength and the muscle fibre cross section of the femoral and gluteal muscles increased by about 40% compared to the control group performing positive (concentric) work at the same heart rate (Lindstedt et al. 2001). Similarly, after six weeks of exercise on the device, the maximum jump height for basketball players was increased by 8% compared to a conventional strength training control group, who showed no growth in this respect (Lindstedt et al. 2001).
Interestingly, the oxygen use of eccentric effort at the same workload is only a sixth or seventh of that at concentric work; that is, the eccentric effort not only results in greater strength, but does so with lower metabolic expenditure (Bigland-Ritchie, Woods 1976).
According to an extensive meta-analysis, it can be said that eccentric exercises are more effective both in building muscle mass (i) and increasing muscle strength (ii); however, since the results achieved in this manner depend closely on the speed and intensity of the exercise, it is not completely clear the extent to which the strength of muscles built and strengthened by eccentric work can be transferred to other movements (Roig et al. 2009).
Eccentric muscle function also has another very important feature, which is the ability to recover elastic energy. For example, while running, when the foot touches the ground, some of the kinetic energy is absorbed by the braking muscles, and then returned on recoiling to the now shortening, contracting muscles. In this case, the musculo-tendon system acts as a kind of spring, which recycles temporarily absorbed energy into the motion, thereby making our movement more energy efficient (i), and also protecting the joints (ii) because it acts as a shock absorber (Lindstedt et al. 2001). Of course, if this elastic energy is not used within a small time window, it will transform into heat and be lost (Lindstedt et al. 2001). This would happen if, for example, we stayed on the ground at every step while running, waited a bit, and then we jumped again. During strength training, this type of flexible energy tends to be used by the athlete in a variety of exercises – e.g. push-ups, pull-ups, bench presses, squats, calf raises, etc. – thus, the athletes are “cheating” a little bit. If, for example, we wait a little while at every end point of a push-up, i.e. we do not immediately release or push our body upwards, then this energy-recycling mechanism, that is, the elastic momentum, is not put into practice, which makes it naturally much more difficult to carry out the exercise.
What role does this elastic energy recovery mechanism play in our movement? According to laboratory measurements, a comfortable jumping frequency chosen by volunteers jumping on their own and a forced deceleration to half this speed resulted in a 50% energy consumption difference: this energy had to be recreated by the muscles acting against gravity during the slow jump (Lindstedt et al. 2001). In other words, this means that the correct choice of the frequency of motion can make our energy use more efficient by at least 50%, precisely because of flexible energy recovery.
Speaking of muscle elasticity, we cannot forget the titin molecule that connects the Z-discs within the muscle fibres. According to older ideas, passive tension during muscle function is ensured by tendons, and muscle cells play no real role in this. However, according to present-day views, within the normal physiological limits of elongation, this passive elasticity is mainly due to intracellular titin (along with other proteins), and the tendons will play a role in stretching beyond that extent (Lindstedt et al. 2001).
2.1.2.5. Muscles and bones as levers
From our basic biological studies we can remember that the muscles can be characterized by different roles depending on the direction in which they move in relation to the midline. They are distinguished as follows:
flexor
extensor
adductor
abductor and
rotator roles.
Each movement is often done in groups: muscles that generate movements in a single direction around a given joint are called agonist or synergist muscles. When they have the opposite function around the same joint, they are antagonist muscles. Let us see how muscles with different roles work as skeletal structures that lift, push, pull and rotate loads.
The elements of the skeletal system connected with muscles can be described as levers, and this is particularly evident in the case of the skeletal elements of the limbs – the operation of the levers is simply perceived as limb movements. The lever is a simple machine that consists essentially of a beam (arm) which is shored up at one point. At a given point, we can exert a force on this beam, and by rotating the beam around the pivot point or fulcrum we can lift a load. The so-called one-sided or one-arm lever (Figure 3) is a lever in which the load and the applied force are on the same side of the lever arm. For example, a wheelbarrow, a nutcracker, or a tweezer operates like this. A good example of this in the human skeleton is the mandibular joint with the masseter muscle and the temporal muscle (Figure 4), or the elbow joint with the biceps muscle (Figure 5). The masseter muscle pulls the mandible to the zygomatic arch, the temporal muscle pulls it to the temporal bone; and the biceps moves the radius towards the scapula.
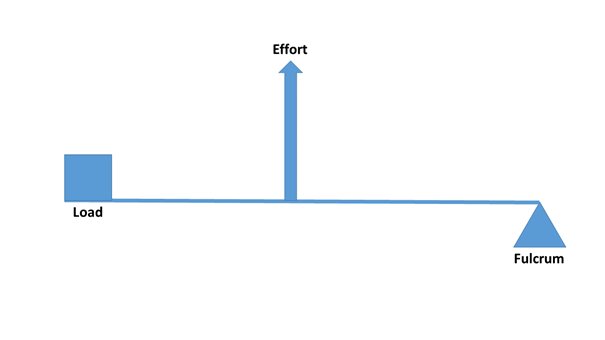
Figure 3: One-sided or one-arm lever(Figure: Author’s own production.)
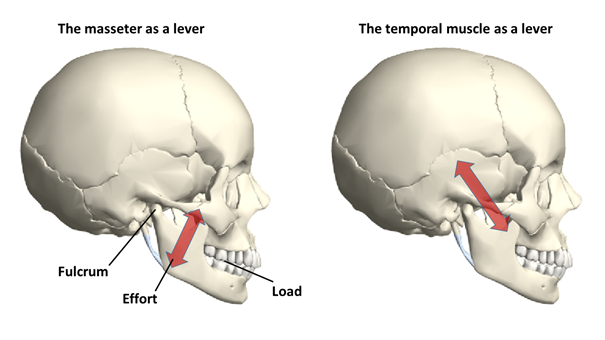
Figure 4: The mandibular joint with the masseter and the temporal muscle acts as a one-arm lever(Source: Wolfram Mathematica.
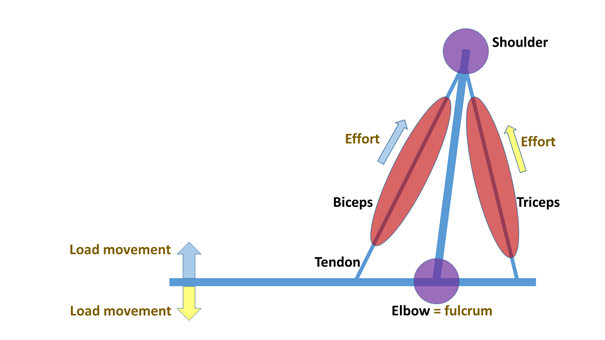
Figure 5: Biceps and triceps as levers(Figure: Author’s own production.)
In a two-sided or two-arm lever (Figure 6), the load and the applied force fall on different sides of the fulcrum. Such levers are scissors, the seesaw, or “traditional” two-arm mechanical scales. In the human body, the triceps muscle also works like this (Figure 5). The triceps brings the back of the ulna to the scapula and the upper arm bone (humerus), thereby distending the arm. It should be noted that the English-language literature traditionally classifies the levers differently, but this type of distinction is of little use to us. [In first-class levers, the fulcrum is centred (e.g. a seesaw, scissors), in second class the load is in the middle (e.g. a wheelbarrow), and in third-class levers, the force is in the middle (e.g. canoeing, tweezers, the human jaw).]
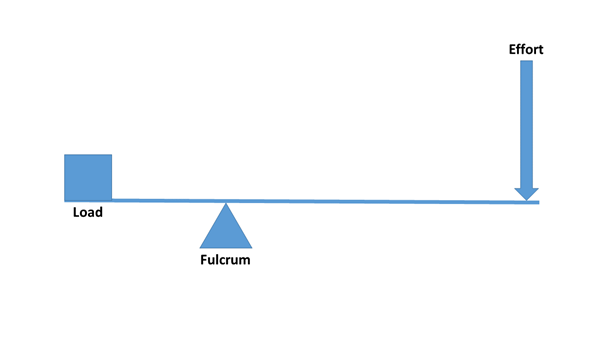
Figure 6: Two-arm lever(Figure: Author’s own production.)
2.1.2.6. Great force required – pennate muscles
The shape and length of the muscles are not unimportant from the point of view of force production. Regarding the relative force of the muscles of different sizes and shapes it can generally be said that shorter and wider muscles are stronger, but they can perform less contraction (i.e. they move something to a shorter distance) than the longer and thinner muscles, of course with the same muscle volume (and output). There are regions in our organism that, due to their layout, require relatively great effort: in such places it is worth using short muscles. However, the problem is that to a given length of bone (and lever arm) only a given length of muscle can be fitted, so as to allow the right amount of force to be applied, while also optimizing the length of the tendons. Thus, it is not possible to “force” a muscle of any length anywhere, even though the magnitude of the effort would justify it. Now, nature has a clever solution to the problem, and it is called the pennate muscle, or more technically, musculus pennatus (Figure 7). In the pennate muscle, a sheet of tendons (an aponeurosis) runs inside the muscle, and muscle fibres are attached to it laterally. Thus, in terms of anatomical arrangement, the optimum size of the muscle can be located within the system, while it can exert greater force through its short muscle fibres than if it were a spindle-shaped muscle. Such a muscle in the human body is, for example, the musculus gastrocnemius (triceps surae or calf; Figure 8), or the musculus triceps brachii (triceps muscle). These muscles have to exert a great deal of strength while lifting the body to the toes, adhering to the heel bone, but even more when running (the calf muscle), or stretching the arm out along the outside of the elbow joint (triceps). Pennate muscles can also be found elsewhere, such as the deltoid muscle, in the thighs, shins, or among the muscles that move the fingers.
The truth is, however, that the pennate muscles also have a drawback in addition to their ability to exert great force. Since the muscle fibres are not parallel to the direction of movement (as in the spindle-shaped muscles), the lateral component of the vector of the contraction force is not utilized during the contraction. This loss is roughly equivalent to the cosine of the angle to the direction of the exerted force – e.g. at a 30 degree angle this means a loss of about 13.4% (Vogel 2013). By the way, the picture is more complex, because during the exertion of force the geometric arrangement of the pennate muscle changes, i.e. the aforementioned angle changes too. In addition, depending on the magnitude of force, the “working profile” of the pennate muscle also changes. It could also be said that the shape transformation of the pennate muscle acts as a kind of automatic transmission, because it tries to increase the speed release at low weight, i.e. in a “high gear”, while in the case of a high weight resistance it increases the force, shifting into a “lower gear” (Azizi et al. 2008). A similar automatic transmission mechanism has been described for the feet and toes, which, due to their structural arrangement, optimize the exertion of force of the extensor muscles that carry out movements around the ankle as a rotation point to the area which stands on the ground, thereby helping to meet the different needs of force transmission between reaching the ground and springing back (Carrier et al. 1994).
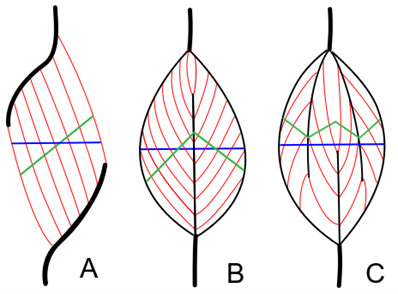
Figure 7: Structure of pennate muscles. A: Unipennate muscle; B: Bipennate muscle; C: Multipennate muscle(Source: Wikipaedia. Uwe Gille - Own work, CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=3181778)
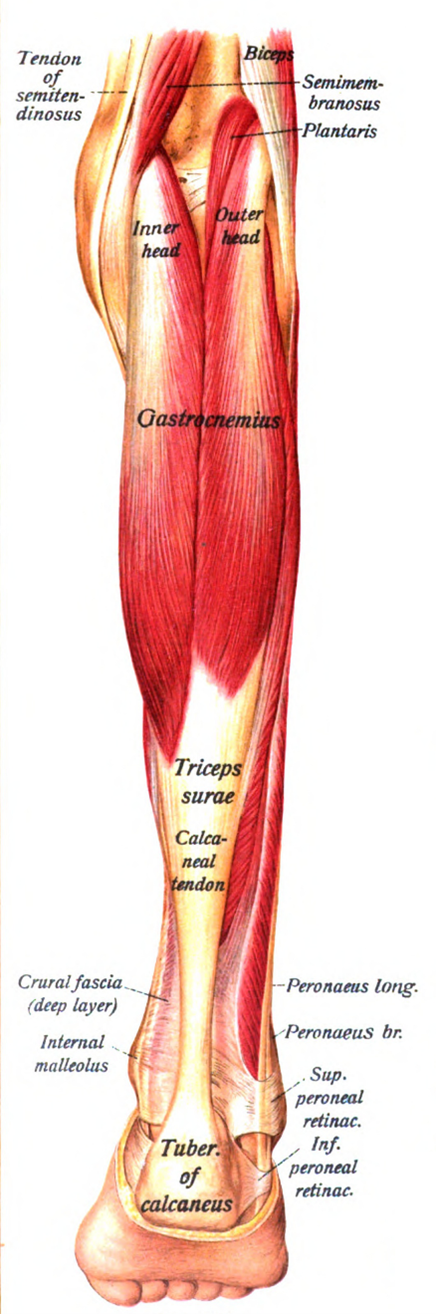
Figure 8: The calf is a bipennate muscle(Source: Wikipaedia. By Dr. Johannes Sobotta - Sobotta’s Atlas and Text-book of Human Anatomy 1909, Public Domain, https://commons.wikimedia.org/w/index.php?curid=29822860)
2.2. The joints
A key factor in sports, especially competitive sports, is to maintain the integrity of the joints. Since this is not easy to achieve with increasing loads, one of the potentially high risks of competition is that the joints become over-used, and unfortunately, often destroyed. Below, we will try to get some insight into the structure of the major joints. In general, it is important to know what the major parts of a joint are, what the tasks for each part are, why and how they may be damaged, and what kind of healing process is faced by those who suffer joint injuries. Here, of course, we only deal with important joints in the field of sports, so, for example, not the suture joints between the skull bones.
The joints with the muscles form important energy transfer elements of the locomotor system. The function of the movable joints is to minimize the motional friction at the joining of the bone ends, and at the same time to ensure the movable connection of the units of the modular limb structure (e.g. upper arm – forearm – hand), and determine the direction of movement of the limb. The articular surface is covered with cartilage (histologically called hyaline cartilage), which protects the bones, reduces friction, and distributes the tension generated in the joint (Figure 9). In addition to the cartilage covering the ends of the bones, in some joints, additional cartilage structures can help with load bearing, such as the cartilaginous rings, formed of fibrocartilage, located in the joint cavity (for example, the menisci of the knee joint or the intervertebral discs between the vertebrae; Figures 10, 16 and 17). Between the cartilage surfaces, joint or synovial fluid provides lubrication, vibration absorption, and thus good mobility. The liquid is located in a space separated by a membrane (synovial membrane) (Figure 9); this membrane and the cartilage cells produce the synovial fluid. From the outside, the articular capsule covers the joint, and outside of this, the joint ligaments running in different, statically and dynamically significant directions ensure the proper attachment of the bones. Fluid-filled flat sacks, called synovial bursae (lat.: bursa synovialis; bursae synoviales) protect vulnerable parts in areas exposed to stronger mechanical impact (under the attachment of ligaments to bones, between joints and muscles) (e.g. Figure 20).
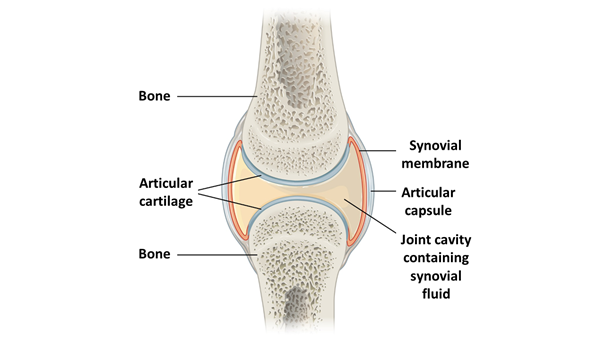
Figure 9: Schematic structure of the joint(Source: Wikipaedia. By OpenStax College - Anatomy & Physiology, Connexions Web site. http://cnx.org/content/col11496/1.6/, Jun 19, 2013., CC BY 3.0, https://commons.wikimedia.org/w/index.php?curid=30131665; adapted)
Due to their function, the articulations between the spinal vertebrae (Figure 10) allow much less room for manoeuvre for the connected vertebrae: they have a much more tightly bounded ligament system compared to the previously described joint structure (it stabilizes them both at each vertebra and across the entire spinal column).
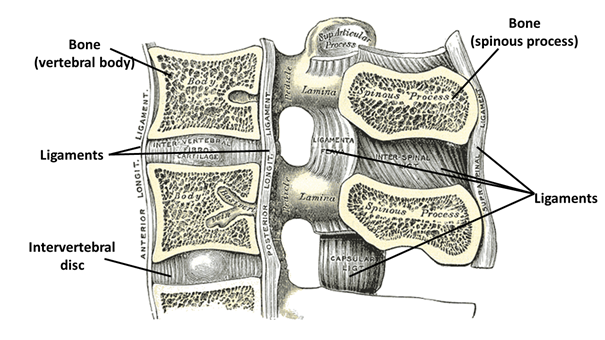
Figure 10: Connection between spinal vertebrae.(Source: Wikipaedia. Prepared by: Henry Vandyke Carter - Henry Gray (1918) Anatomy of the Human Body. Bartleby.com: Gray’s Anatomy, Plate 301, Közkincs, https://commons.wikimedia.org/w/index.php?curid=423008)
2.3. The load responses in the locomotor system
2.3.1. Nerve-muscle harmony: neuromuscular coordination
Nerve cells form an effective signal receiving, processing and transmitting system, while the musculature, made up of muscle cells (and the additional structures) form the effector system of most of the body’s responses. The two, of course, operate in a nested and coordinated manner, so the muscle cells are actually the ‘operating units’ of the nervous system; that is, they are its extensions that execute the motional commands. It is no coincidence that there are two types of cells in our body that can transport the impulse signal generated from the difference of potential formed between the two sides of the cell membrane: the nerve cell and the muscle cell.
The motor information reaches the muscles travelling through the motor nerves that are attached to the muscle cells through the neuromuscular junctions (the acetylcholine is its neurotransmitter). A motor neuron and the muscle fibres that it controls form a functional unit: this is the motor unit. A motor unit consists of one type of fibre (see: Sports Physiology). There are also larger and smaller motor units in the muscle, as a motor neuron can innervate from a dozen to two thousand muscle fibres. At the start of a movement, the Size Principle of motor unit recruitment operates in the muscle; that is, the smallest units turn on first (the stimulus threshold of their motoneurons is lower than that of their larger counterparts), and then the larger units follow. The largest motor units are rarely used, as they are mobilized with a sudden, great effort (accordingly, they are usually composed of a high-strength but soon exhausted cell type).
In most sports, it is necessary to increase physical strength, which in some sports can even involve targeting extreme ranges. For example, one of the great ways to increase strength is to increase the coordination of the motor units of the muscles involved in the given movement. One of the consequences of this is that the number of motor units involved in the movement increases, which naturally allows the generation of greater force (i) (Katics, Lőrinczy 2010; Radák 2016). This type of increase in strength is particularly useful for participants in weight competitions, as it is very important for them to be able to gain strength without increasing muscle mass (and thus body weight). The alignment of the motor units also entails the refinement of the movement (ii), thus increasing the efficiency of any type of movement. Therefore, this kind of movement development – from carrying out graceful gymnastics movements, to subtle, deceptive fencing moves or other combat movements, right through to complex ice dance choreography – can be exploited in every field.
But why is this possible at all? Why and how does the muscular system develop beyond quantitative growth, into qualitative growth, too? This adaptation is made possible by a very important ability of our nervous system, which is called, in short, neuroplasticity. Our nerve cells form a networked system whose task is to respond ever more effectively to the external or internal effects (stimuli) that affect us. Naturally, as these incoming effects change, our nervous system also tries to follow them, i.e. it continuously optimizes the functions of signal processing and response. The cellular basis of this optimization is the phenomenon that the connection-seeking projections (dendrites) of the neurons are constantly moving at a demonstrably significant rate already on a time scale of ten to twenty seconds (Fischer et al. 1998, Matus 2000), and are looking for new connections. If signalling occurs at the new junction point, the synapse is strengthened, i.e., molecular changes in the membranes of the two cells that exchange information make the signal transfer more efficient. Conversely, less frequently used information paths will be degraded. This massive synaptic strengthening and weakening results in neural networks with different operating patterns than before; in other words, the nervous system itself changes in response to experience. And since the nervous system is closely connected to the muscular system, the functioning of the muscles also adapts to this process. Thereby, neural optimization also means muscular optimization: our movement becomes more refined and more coordinated. In addition, the muscle may undergo volumetric changes as well as circulatory and biochemical adaptation – but these topics will be dealt with in the physiological section.
2.3.2. Factors affecting the mechanical properties of bones and joints
2.3.2.1. Immobilization and disuse
The possible and actual responses to loadings of our tissues are significantly influenced by certain circumstances and characteristics. One of the most important of these is immobilization and non-use.
With shorter or longer periods of forced rest in plaster, we may have experienced the well-known phenomenon that during a few weeks of non-use, muscle mass and strength are significantly reduced. However, it is perhaps less well known that the carrying capacity of the joint ligaments is also proportionally reduced, which makes the joint weaker, stiffer and more vulnerable. In addition, bone mass is also decreasing, which also contributes to increasing vulnerability (Bartlett 2005). In immobilisation periods of 9 to 12 weeks during animal experiments, the strength of various tested joints decreased by as much as 30-60% (Loitz, Frank 1993; Bartlett 2005).
It is also important to know that the loss of connective tissue caused by forced rest occurs much more quickly than rebuilding it to the previous level. The load-bearing capacity of the anterior cruciate ligament in the knee joint of monkeys decreased by about 39% after eight weeks in plaster, the recovery of the previous load-bearing capacity was only partial even after 20 weeks, and even after regular practice it took almost one year to reach a level of 91% (Noyes et al. 1974, Gabriel 2002). Interestingly – but logically – isotonic muscle tension exercises during plaster application had a positive effect on the muscles, but had no effect on the condition of the ligaments: their condition deteriorated during non-use (Noyes et al. 1974). So, if it is necessary to pause a given sports activity, and immobilize the particular limb, it is advisable to carry out some kind of cautious, level-maintaining training, if the circumstances allow.
2.3.2.2. Age and sex
Of course, age and sex are also important factors influencing the mechanical properties of tissues. The growth occurring during adolescence is of paramount importance, because during this period the increase in the weight and density of the bone mass is significantly accelerated; this is also accompanied by changes in muscles (even if, usually, at a slower pace), and body proportions undergo transformation. In this way, the structure, weight distribution and balance characteristics of the locomotor system change, and this naturally also affects the movement properties of the body.
And with the advent of old age, the weight of the skeleton is reduced, the muscles atrophy, and the connective tissue (tendons, ligaments) is also weakened, which makes it impossible to practice sport at the previous intensity. It should be emphasized, however, that the body reacts to the mechanical effects with a high degree of adaptation, so life-long sporting activity has a significant protective effect on the integrity of the locomotor system. Therefore, those who start sport at a greater age are much more vulnerable than those who have also done sport previously, so at an older age beginners – and their trainers – need to pay more attention to workouts.
As the body ages, the slow degradation of the trabecular (spongy, cancellous) bone is a natural process. In the trabecular system, disintegration foci are formed, from which the cancellous bone gradually and partially disappears, thereby reducing the carrying capacity of the bone. This process is most pronounced in regions with a significant proportion of the spongy tissue in the bone volume and which are subjected to significant mechanical stress, such as in the body of the spinal vertebrae or the femoral neck. It is no coincidence that fractures often occur in these two zones during a fall in old age. The process of age-related bone reduction is more pronounced in women, as menopause results in decreased levels of female hormones in the body. These hormones have previously had a sort of protective effect on bone tissue (oestrogens inhibit the activity of osteoclast cells that break down bone tissue, so that the balance of remodelling in the bone tissue is shifted towards the demolition, as a result of the decrease in female hormones levels).
As stated above, the effect of age is also present in connective tissues. According to some studies, the tensile strength of the anterior cruciate ligaments of the knee joint decreases to an average of about one third from a young age to the age of 65. However, this deterioration can be reduced by about 50% by regular exercise (Bartlett 2005). In terms of gender differences, it should be emphasized that the maximum bone mass and bone density values in women are on average lower than in men. The stronger musculoskeletal system that develops as a result of the male hormonal background would create unequal chances for gender rivalry in most sports. However, in contact sports, the difference in skeletal capacity is another reason why boys and girls compete separately after a certain age, as the risk of injury to girls/women would be disproportionately increased.
2.3.2.3. Exercise, training
Regular practice has the opposite effect to disuse and immobilization. Therefore, physically trained people naturally have less chance of having sports injuries. Compressive forces and tensile forces exerted by the musculature in the normal range stimulate bone formation, both locally and systemically. In addition to increased muscle mass, bone density, bone mass, and connective tissue strengthening, the surface of the bones also changes, as the protrusions on the bone surfaces that that serve as attachment sites for tendons, also grow. Thus, in athletes, the crests, spurs, and condyles of the bone surface appear more articulate than they do in those who do not perform physical activity. Of course, the changes also take place deep inside the bone, and not only on the surface: we can remember that the trabecular tissue has a lower mineral content and faster remodelling capacity than the compact tissue; the remodelling rate is about eight times that of the cortical bone tissue (25% and 3% per annum; Manolagas, Jilka 1995). The bone trabeculae found here take on a new spatial arrangement due to changing forces. On the other hand, if the pressure on the bones is extremely high for a long time, the bone tissue will atrophy (Bartlett 2005).
In addition to the bones, the joints naturally also show adaptability over the long term. The substance of tendons and ligaments gains in volume and mass, and their carrying capacity also increases (Bartlett 2005). However, in addition to the strengthening of the stabilizing ligament system, in the case of regular over-stretching, the range of movement of the joints may also be altered if this is the objective (for example, doing the splits, bridges, extreme bending of the spine, etc.). However, it is important to know that the expansion of the movement range also increases the chance of injury to the particular joint, since the joint’s ligament system is no longer able to perform its stabilizing function to the former degree.
Within its limits of flexibility, cartilage tissue also adapts to physical activity: it increases both its thickness and elasticity (Bartlett 2005).
2.3.3. Short-term adaptation: warm-up
During physical activity, changes occur in the joints in the short and long term. The short-term change in the joints is mainly an increase in the quantity and viscosity of the synovial fluid. When motion activity increases – for example, when we warm up – then the fluid production of the synovial membrane cells increases. If the load is high, the viscosity (i.e. the internal friction) of the joint fluid produced will also increase, thereby allowing the joint to withstand higher mechanical effects. At this point, for example, we can immediately find a very important reason for the importance of proper warming up. Unfortunately, in many training sessions, especially in sports which do not have a background of an extensive competitive structure and appropriate training for coaches, trainers “save” on at least part of the warm-up because they are not aware either of the use of a good warm-up or how to structure it. In addition, of course, athletes themselves, especially hobby athletes, also try to play around during the coach’s warm-up, e.g. they blow their nose for a long time, they always tie their shoelaces, go to the bathroom during the warm up and stay there for an unreasonably long time, or arrive late and then do not warm up independently to a sufficient degree, etc. If the body then gets a heavier load, some joints may become dislocated and their ligaments may be strained; and in the long run, they have to deal with chronic arthritis, and problems caused by worn joints.
The increased body temperature also increases the responsiveness of the muscles: the speed of contraction increases, the time to reach peak tension decreases, and the value of the power delivered increases (Bartlett 2005). Appropriate warming-up therefore has two benefits: protecting from injury and increasing the performance achieved in training or competition.
2.3.4. Response to overload: injury
Unfortunately, injuries are a part of sports. Injury occurs when a biomechanical force of such direction and magnitude is exerted on a given tissue or organ that it exceeds its carrying capacity. Injuries can occur as a result of a single, acute trauma, or may result from recurrent chronic overload.
While sports practiced to a rational extent effectively increase the overall fitness of the body and hence its ability to protect itself against injuries, sports activities – especially competitive sports – simultaneously also increase the risk of injury. One reason for this is that if there is a greater amount of movement (especially movements carried out in ranges with a high risk of injury), there is a higher chance of injuries caused by inappropriate movements or accidents. Another reason is of a simple physiological nature. During sports, we can enter a state in which the heightened stress response (as the physical stress also triggers a stress response) increases our pain sensation threshold significantly. The evolutionary explanation for this is that in this way, escape from a dangerous situation is not hindered by the appearance of a sensation of pain. However, we know that nociception is a system that indicates to the brain that we have left the optimum range for certain parameters, and that the given situation carries the risk of tissue damage. Because of this, an unpleasant pain sensation appears that encourages the person to eliminate the situation (that’s why the nociceptors do not adapt to the stimulus that triggers the pain, but maintain their firing during the whole stimulation, or even increase it). In this way, however, during sports, this signalling system within the body does not always work properly, which also risks causing the damage not to be noticed in time or be given due attention. After sports, with a cooled down body and normalized nociceptive thresholds, of course, the pain breaks through, and the previously ignored injury can be felt to its full extent.
It is important to know that the risk of injury is greatly increased by
insufficient warm-ups,
fatigue and
overtraining.
These are all factors that knowledgeable athletes can control themselves relatively easily, and thus already eliminate most of the risk factors leading to potential injury.
In terms of injuries, the adolescent growth explosion and old age are more sensitive periods, and contact sports are high risk, and it is known that in team sports there are more injuries during contests and matches than during training (Bartlett 2005).
Approximately 90% of sports injuries are strains or shock-induced contusions (Järvinen et al., 2005, 2014), and mostly affect joints and their related soft tissues, and only to a lesser extent bones (Bartlett 2005). Below, we discuss the main types of injuries.